Issue №1. Love and science
Our first issue is devoted to the famous mysterious feeling - Love. We have collected articles describing scientific views on it.
Study the articles using Explore button at the top-right corner of the page.
Article №1
When love and science double date
Sure, your heart thumps, but let’s look at what’s happening physically and psychologically
February 13, 2018
“They gave each other a smile with a future in it.”
—Ring Lardner
Love’s warm squishiness seems a thing far removed from the cold, hard reality of science. Yet the two do meet, whether in lab tests for surging hormones or in austere chambers where MRI scanners noisily thunk and peer into brains that ignite at glimpses of their soulmates.
When it comes to thinking deeply about love, poets, philosophers, and even high school boys gazing dreamily at girls two rows over have a significant head start on science. But the field is gamely racing to catch up.
One database of scientific publications turns up more than 6,600 pages of results in a search for the word “love”. The National Institutes of Health (NIH) is conducting 18 clinical trials on it (though, like love itself, NIH’s “love” can have layered meanings, including as an acronym for a study of Crohn’s disease). Though not normally considered an intestinal ailment, love is often described as an illness, and the smitten as lovesick. Comedian George Burns once described love as something like a backache: “It doesn’t show up on X-rays, but you know it’s there.”
Richard Schwartz, associate professor of psychiatry at Harvard Medical School (HMS) and a consultant to McLean and Massachusetts General (MGH) hospitals, says it’s never been proven that love makes you physically sick, though it does raise levels of cortisol, a stress hormone that has been shown to suppress immune function.
Love also turns on the neurotransmitter dopamine, which is known to stimulate the brain’s pleasure centers. Couple that with a drop in levels of serotonin — which adds a dash of obsession — and you have the crazy, pleasing, stupefied, urgent love of infatuation.
It’s also true, Schwartz said, that like the moon — a trigger of its own legendary form of madness — love has its phases.
“It’s fairly complex, and we only know a little about it,” Schwartz said. “There are different phases and moods of love. The early phase of love is quite different” from later phases.
During the first love-year, serotonin levels gradually return to normal, and the “stupid” and “obsessive” aspects of the condition moderate. That period is followed by increases in the hormone oxytocin, a neurotransmitter associated with a calmer, more mature form of love. The oxytocin helps cement bonds, raise immune function, and begin to confer the health benefits found in married couples, who tend to live longer, have fewer strokes and heart attacks, be less depressed, and have higher survival rates from major surgery and cancer.
Schwartz has built a career around studying the love, hate, indifference, and other emotions that mark our complex relationships. And, though science is learning more in the lab than ever before, he said he still has learned far more counseling couples. His wife and sometime collaborator, Jacqueline Olds, also an associate professor of psychiatry at HMS and a consultant to McLean and MGH, agrees.

More knowledge, but struggling to understand
“I think we know a lot more scientifically about love and the brain than we did a couple of decades ago, but I don’t think it tells us very much that we didn’t already know about love,” Schwartz said. “It’s kind of interesting, it’s kind of fun [to study]. But do we think that makes us better at love, or helping people with love? Probably not much.”
Love and companionship have made indelible marks on Schwartz and Olds. Though they have separate careers, they’re separate together, working from discrete offices across the hall from each other in their stately Cambridge home. Each has a professional practice and independently trains psychiatry students, but they’ve also collaborated on two books about loneliness and one on marriage. Their own union has lasted 39 years, and they raised two children.
“I think we know a lot more scientifically about love and the brain than we did a couple of decades ago … But do we think that makes us better at love, or helping people with love? Probably not much.”
—Richard Schwartz, associate professor of psychiatry, Harvard Medical School
“I have learned much more from doing couples therapy, and being in a couple’s relationship” than from science, Olds said. “But every now and again, something like the fMRI or chemical studies can help you make the point better. If you say to somebody, ‘I think you’re doing this, and it’s terrible for a relationship,’ they may not pay attention. If you say, ‘It’s corrosive, and it’s causing your cortisol to go way up,’ then they really sit up and listen.”
A side benefit is that examining other couples’ trials and tribulations has helped their own relationship over the inevitable rocky bumps, Olds said.
“To some extent, being a psychiatrist allows you a privileged window into other people’s triumphs and mistakes,” Olds said. “And because you get to learn from them as they learn from you, when you work with somebody 10 years older than you, you learn what mistakes 10 years down the line might be.”
People have written for centuries about love shifting from passionate to companionate, something Schwartz called “both a good and a sad thing.” Different couples experience that shift differently. While the passion fades for some, others keep its flames burning, while still others are able to rekindle the fires.
“You have a tidal-like motion of closeness and drifting apart, closeness and drifting apart,” Olds said. “And you have to have one person have a ‘distance alarm’ to notice the drifting apart so there can be a reconnection … One could say that in the couples who are most successful at keeping their relationship alive over the years, there’s an element of companionate love and an element of passionate love. And those each get reawakened in that drifting back and forth, the ebb and flow of lasting relationships.”
Children as the biggest stressor
Children remain the biggest stressor on relationships, Olds said, adding that it seems a particular problem these days. Young parents feel pressure to raise kids perfectly, even at the risk of their own relationships. Kids are a constant presence for parents. The days when child care consisted of the instruction “Go play outside” while mom and dad reconnected over cocktails are largely gone.
When not hovering over children, America’s workaholic culture, coupled with technology’s 24/7 intrusiveness, can make it hard for partners to pay attention to each other in the evenings and even on weekends. It is a problem that Olds sees even in environments that ought to know better, such as psychiatry residency programs.
“There are all these sweet young doctors who are trying to have families while they’re in residency,” Olds said. “And the residencies work them so hard there’s barely time for their relationship or having children or taking care of children. So, we’re always trying to balance the fact that, in psychiatry, we stand for psychological good health, but [in] the residency we run, sometimes we don’t practice everything we preach.”
“There is too much pressure … on what a romantic partner should be. They should be your best friend, they should be your lover, they should be your closest relative, they should be your work partner, they should be the co-parent, your athletic partner. … Of course everybody isn’t able to quite live up to it.”
—Jacqueline Olds, associate professor of psychiatry, Harvard Medical School
All this busy-ness has affected non-romantic relationships too, which has a ripple effect on the romantic ones, Olds said. A respected national social survey has shown that in recent years people have gone from having three close friends to two, with one of those their romantic partner.
“Often when you scratch the surface … the second [friend] lives 3,000 miles away, and you can’t talk to them on the phone because they’re on a different time schedule,” Olds said. “There is too much pressure, from my point of view, on what a romantic partner should be. They should be your best friend, they should be your lover, they should be your closest relative, they should be your work partner, they should be the co-parent, your athletic partner. There’s just so much pressure on the role of spouse that of course everybody isn’t able to quite live up to it.”
Since the rising challenges of modern life aren’t going to change soon, Schwartz and Olds said couples should try to adopt ways to fortify their relationships for life’s long haul. For instance, couples benefit from shared goals and activities, which will help pull them along a shared life path, Schwartz said.
“You’re not going to get to 40 years by gazing into each other’s eyes,” Schwartz said. “I think the fact that we’ve worked on things together has woven us together more, in good ways.”
Maintain curiosity about your partner
Also important is retaining a genuine sense of curiosity about your partner, fostered both by time apart to have separate experiences, and by time together, just as a couple, to share those experiences. Schwartz cited a study by Robert Waldinger, clinical professor of psychiatry at MGH and HMS, in which couples watched videos of themselves arguing. Afterwards, each person was asked what the partner was thinking. The longer they had been together, the worse they actually were at guessing, in part because they thought they already knew.
“What keeps love alive is being able to recognize that you don’t really know your partner perfectly and still being curious and still be exploring,” Schwartz said. “Which means, in addition to being sure you have enough time and involvement with each other — that that time isn’t stolen — making sure you have enough separateness that you can be an object of curiosity for the other person.”
Article №2
The neuroendocrinology of love
Abstract
Romantic love could be considered as a collection of activities associated with the acquisition and retention of emotions needed to survive and reproduce. These emotions change the individual's behavioural strategies in a way that will increase the likelihood of achieving these goals. love may be defined as an emergent property of an ancient cocktail of neuropeptides and neurotransmitters. It appears that lust, attachment and attraction appear to be distinct but intertwined processes in the brain each mediated by its own neurotransmitters and circuits. These circuits feed on and reinforce each other. Sexual craving is mediated by testosterone and oestrogen and has the amygdala as an important centre. Attraction is mediated by hormones of stress and reward including dopamine, norepinephrine cortisol and the serotinergic system and has the nucleus accumbens the ventral tegmental area as key mediators.
Keywords: love, monogamy, neuroendocrine, oxytocin, prairie vole, vasopressin
He will not know what all but he do know.
And as he errs, doting on Hermia's eyes.
So I, admiring of his qualities.
Things base and vile, holding no quantity.
love can transpose to form and dignity.
love looks not with the eyes, but with the mind.
And therefore is winged Cupid painted blind.
Nor hath love's mind of any judgment taste.
Wings and no eyes figure unheedy haste.
—William Shakespeare
Midsummer Night's dream (1.1.232-243)
Introduction
From an evolutionary perspective romantic love could be considered as a collection of activities associated with the acquisition and retention of emotions needed to survive and reproduce. These emotions change the individual's behavioral strategies in a way that will increase the likelihood of achieving these goals.[1] The enduring question for science has been that if these evolutionarily determined behaviors have a biologic substrate and correlation with activation of specific brain areas (and hormones)?[2] This review attempts to summarize our current understanding of the neuroendocrinology of romantic love.
A Definition of Love
While poets and philosophers are more adept at defining love, for the purposes of this review, this particular definition seems to be apt: love is an emergent property of an ancient cocktail of neuropeptides and neurotransmitters.[3]
The Dichotomy Between Courtship and Sex
Despite the intimate intertwining of the sexual drive with courtship, for over 4 decades investigators have suggested that these processes may be distinct [4] but operate in tandem [Table 1]. Human romantic love (hereafter just love) is cross-cultural, universal, and associated with distinct physiologic, psychological, and behavioral traits.[5] Many of these traits are also characteristic of mammalian courtship which includes increased energy, focused attention, obsessive following, affiliative gestures, possessive mate guarding, goal-oriented behavior, and motivation to win a preferred partner.[2]
Table 1
Comparison of three motivations involved in “love”
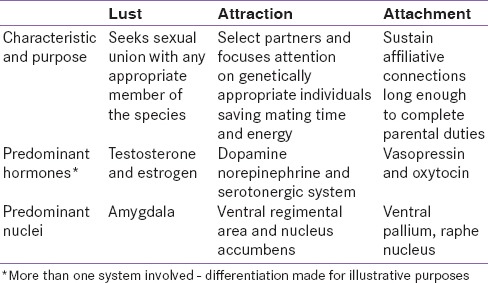
In humans, love often begins when an individual starts to regard another individual as special and unique. This is followed by focused attention, aggrandizement of traits and worth of the object of attention, and minimizing of his or her faults. There is increased ecstasy when things go well, despair when they do not and separation anxiety when apart.[6] Emotional dependence, empathy, sacrifice, and obsessive thinking are common. Sexual desire, intense sexual possessiveness, and mate guarding are present, but the emotional union appears to supersede the craving for sex. Rejection triggers protest and rage, moving into resignation and despair. It has been suggested that love as a preference system is associated with action and conditioning of specific neuroendocrine pathways.
Love as a Primordial Drive
It has been proposed the love is not primarily an emotion but a motivation system (i.e., a system oriented around the planning and pursuit of a specific want or need) designed to enable suitors to build and maintain an intimate relationship with a specific mating partner.[7] Functional magnetic resonance imaging (MRI) studies demonstrate the involvement of areas associated with motivation and goal-oriented behavior in love suggesting that love is a primary motivation system a fundamental human mating drive.[8] Several lines of evidence support this view that love is not an emotion but motivation and are reviewed elsewhere. (8) love also appears to be stronger than sex drive-those rejected by sexual overtures rarely kill themselves or others. Abandoned lovers sometimes stalk, commit suicide homicide, or fall into clinical depression.[7]
Stress as an Initiator/Facilitator of love
The early phase of love represents an extreme neurobiological state somewhat contradictory in a physiologic sense from subsequent phases and states. Stress appears to be the trigger for a quest for pleasure, proximity, and closeness. As a norm, moderate stress encourages social interaction.
Within a homeostatic range, stress-related physiologic processes including the hormones of the hypothalamo-pituitary adrenal axis can help develop and promote social bonding.[9] Indeed, some of the signs commonly associated with love-anxiety palpitations increased peristalsis are manifestations of the stress response (albeit in a pleasurable way). Indeed, subjects in love show higher levels of cortisol when compared to controls.[10] This “love induced hypercortisolemia” may represent a nonspecific stress response to change that characterizes early phases of relationships or a physiologic state of alertness that may help overcome neophobia. Irrespective, this stress response appears to be important in the formation of social contact and attachment.
Central norepinephrine may also be involved. Increased activity of norepinephrine generally produces alertness energy sleeplessness and loss of appetite, increased attention, and increased memory for new stimuli which characterize (the earlier) phases of human love.[11] Norepinephrine is also associated with peripheral sympathetic nervous systems including increased heart rate, sweating trembling which may explain this experience in love.[12]
Positive social interactions and pair bonding (see below) appear to alleviate stress through oxytocin(OT) facilitating security and support. It appears therefore that initial anxiety and stress is an inherent component of early love which reaches its fulfillment through “the chill” rendered by love and deep relationships. These appear to be mediated by a complex interaction between pathways that link stress response to reward mechanisms. Indeed, serotonin-dependent pathways such as the amygdala appear to interact with OT (see below). OT administration appears to decrease anxiety by inhibiting amygdala activity.[13]
Gonadal hormones
The role of gonadal hormones in this regard appears to be facilitatory but peripheral in love. Sex hormones may exert developmental effects on neural systems involved in social attachments and may mediate both genetic and environmental influences on the propensity to love and form attachments.[14] Testosterone receptors are distributed in the hypothalamus. Testosterone through these receptors may suppress levels or activity of serotonin which apparently increases aggressiveness. Testosterone further enhances vasopressin levels in the medial amygdala lateral hypothalamus and pre-optical medial area which are involved in aggressive behaviors.[15] Gonadal hormones may further regulate OT and vasopressin through indirect mechanisms. However, social attachment does occur even in the absence of gonadal steroids suggesting that gonadal hormones are only a small piece of an intricately knit puzzle that form the complex phenomenon called love.
Gender differences evident on functional imaging in the partner preference and early phases of love warrant mention. Men show more activity in a region of the right posterior dorsal insula (an area correlating with penile turgidity and viewing of beautiful faces) and in regions associated with the integration of visual stimuli. Women tend to show more activity than men in regions associated with attention, memory, and emotion. Courting men respond more strongly than women to visual signals of youth and beauty. Women appear to be more attracted to men who offer status and resources.[16]
Vasopressin oxytocin and “Pair Bonding”
Pair bonding is a very bland scientific term for enduring (romantic) relationships (attachment) and is seen in >5% of the mammalian species.[17] Pair bonding across species is defined as an enduring preferential association formed between two sexually mature adults and is characterized by selective contact, affiliation, and copulation with the partner over a stranger.[18] These are associated with other complex behaviors including mate guarding and biparental care of the young.
Pair bonding evolved most likely, as an adaptive response to the need of additional parental investment in the rearing of the young and mechanisms through which this relationship was preserved (mate guarding). In other words, romantic relationships and their persistence (through monogamy) was an evolutionary necessity in species in which bi-parental care of the offspring was critical. There are clear benefits to both partners of the relationships as well. In humans, individuals in stable marital relationships live longer than single individuals cross demographic groups. High levels of intimacy correlate negatively with depression and positively with immune function and cardiovascular health.[19]
Most of our knowledge on the neuroendocrinology of intimacy is based on the work on the prairie vole (Microtus ochrogaster) a humble but socially monogamous model from the grasslands of the central United States. In this harsh grassland with scarce resources, the prairie vole evolved into a monogamous animal with breeding pairs living together until one partner dies; the surviving partner does not find another mate. The male prairie vole is highly paternal, helps with nest building guards the nest from conspecific strangers. In general, an animal with low levels of aggression, the male displays enhanced levels of aggression toward strange males. It displays high levels of paternal behavior to litters; this extends to juveniles even after a second litter is born.
Arginine vasopressin (AVP) OT and dopamine (DA) have been reported to be important in regulating social behavior including sexual behavior, aggression, and maternal care. While there are no differences in AVP and OT neurons or their distribution between the monogamous prairie voles and their polygamous cousins, remarkable differences are seen in the receptor distribution (the V1aR and OT receptor [OTR]) and their densities. Interestingly, these densities are stable across the lifespan of the vole. These differences may explain by subtle differences in the promoter region the V1aR and the OTR.[20] The human version of this gene has similar polymorphisms. It is possible that epigenetic modifications of the OTR are also involved.[21]
In the male prairie vole, cohabitation with mating appears to increase AVP synthesis in the bed nucleus of the stria terminalis and AVP release in the limbic system. In the female chemosensory clues altered OTR density in the AOB. Activation of the OT and vasopressin receptors in these centers might result in the development of a conditioned partner preference in prairie voles. Antagonism of the OTR impairs the formation of pair bonds. This effect appears to be larger in the female.
Knockdown of VP production in the paraventricular nucleus of the hypothalamus in the zebra finch increases aggressiveness in the male decreases it the female and reduces gregariousness in both. Knockdown of OT reduces gregariousness, pair bonding, nest cup ownership and side by side perching in females and induces hyperphagia in males.[22] An association between arginine vasopressin receptor 1A polymorphism and human pair bonding behavior analogous to voles has been reported.
AVP activity in the ventral palladium affects partner preference. V1aR activation in this region is necessary for pair bond formation.[23] Similarly, activation of the OT in the nucleus accumbent (NA) also contributes to partner preference and pair bonding.
Functional MRI studies of human partners in long-term relationships show activation the ventral palladium putamen the anterior cingulate cortex and the mid-insular cortex.[24] The ventral putamen/palladium region in particular corresponds to the distribution of V1a receptors in the prairie vole. The areas appear, and connections appear to be distinct but related to those for maternal love. Maternal love activated specific different areas including the lateral orbit frontal cortex but also some same areas as (romantic) love including medial insula, the anterior cingulate gyrus, and caudate nucleus. Both appear to share areas rich in OT and AVP receptors.
Love as a Reward
From the very beginning of our efforts, in understanding the biologic basis of love it has been clear that it involves reward centers in the brains. In this love and addictions (such as by drugs) are somewhat interconnected the one key difference is that naturally rewarding activities such as love are controlled by feedback mechanisms that activate aversive centers that limit the destructive qualities of addiction seen with drugs.[25] Love activates specific regions in the reward system. The effects include a reduction in emotional judgment and reduced fear and also reduced depression and enhanced mood. It also leads to a reduced need to assess the social validity of that person.[26] It thus appears to deactivate areas mediating negative emotions, avoidance behavior critical social assessment and, on the other hand, triggers mechanisms involved in pleasure reward and appetitive motivation [Figure 1].[27]
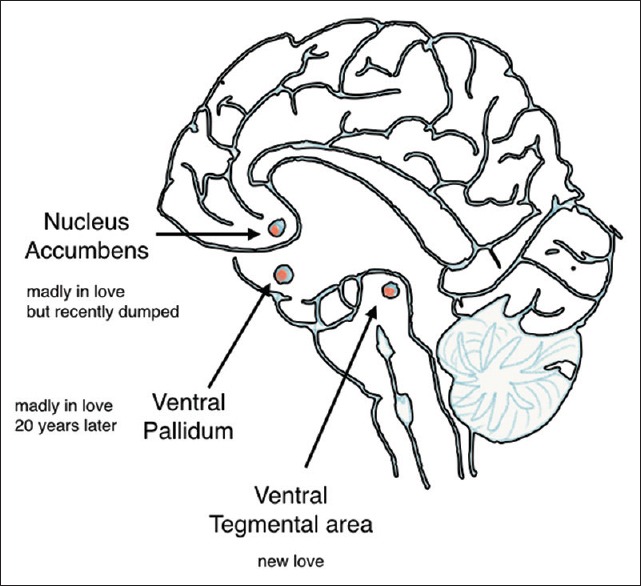
Nuclei associated with motivations collectively involved in “love”
Studies which have examined the OT and AVP receptors strongly suggest that the activation of these receptors in the reward circuitry is important for the development of pair bonding. As a critical part of the reward process, DA appears to be central to the maintenance of love. Differences in DA and its receptor distribution densities have been reported in vole studies (vide above). Dopaminergic pathways appear to be more specific for partner preference that attachment.[28]
While several DA systems exist in the brain, the mesolimbic DA system appears to be the most important in this respect. Both D1 and D2 receptors though partially functional antagonists are both significantly expressed in the NA. Other DA receptors (D3-5) are also linked to the limbic system and are substantially present in the amygdala and the hippocampus. Their functions include reward and motivation and appear to share common morphologic evolutionary and molecular roots. Endogenous opioids may also be involved in this process.
Early studies that involved functional MRI, which used the partner's photograph as a visual stimuli confirm the involvement of the right ventral segmental area (VTA) which is a central region of the brain's reward system [29] associated with pleasure, general arousal, focused attention, and motivation to pursue and acquire rewards.[24] The VTA projects into several regions including the caudate nucleus which plays a role in reward detection, expectation, representation of goals, and integration of sensory inputs to prepare for action. These appear to be true of both early intense (7.4 months) and a little later and not so intense love (28.8 months).[30]
The Role of Sexual Activity
Clearly, sexual activity is an important component of the reinforcement of the reward system, and this appears to reinforce attachment. Increasing levels of testosterone and oestrogen promote DA release.[31] Similarly, elevated activity of dopaminergic pathways appear to increase the release of testosterone and oestrogen. The relationship between elevated central DA sex steroids, sexual arousal, and performance appears to be conserved in humans (7). The sympathetic nervous system also appears to contribute.
Behavioral data support the complementary but distinct pathways for love and sex drive: (a) While sexual drive is often expressed toward a range of individuals while love is focused on one particular individual (b) the sex drive can be quelled when satiated; love does not decrease with coitus and persists unabated for months or years. Sex drive enables individuals to initiate courtship and mating with a range of partners; love focuses mating energy to specific individuals conserving time and metabolic energy (6).
Pleasure and reward activate behavioral patterns that get memorized for the goal of repetition and faster and better recognition later. There is clear evidence to support a connection between attachment behaviors and pleasure pathways that involve hippocampal mechanisms.
The VP is a major target of the NA. The interaction of OT, AVP, and the DA systems within the reward circuitry appears to be the foundation of monogamy. It is hypothesized that in monogamous species (such as the prairie vole) sex triggers the activity of AVP in the ventral pallidum and OT in the NA and facilitates DA release in these reward regions which in turn motivates the male and female to prefer a current mating partner and initiates attachment or pair bonding. In promiscuous species, the male feels the attraction but does not associate the pleasurable feeling with a specific partner and so does not initiate long-term attachment (23). The relationship with the DA reward systems also appears weaker in these species.[32] Complex interactions between gonadal reward and sympathetic systems demonstrate there are distinct but overlapping neural networks involved in love and sex the latter contributing to the reinforcement of the former. Their interdependence also distinguishes romantic love from more platonic attachments including friendship and maternal love (vide above).
It is interesting to extrapolate this to humans. Humans engage in sexual activity throughout the cycle which may serve to strengthen the pair bond. Interestingly and in contrast to other species human females have enlarged mammary tissue independent of lactation. Breast and nipple stimulation are integral to human sexual activity.[33] Nipple stimulation during lactation is one of the most potent stimulus for OT release.[34] This part of sexual activity may reinforce pair bonding in humans. OT levels are elevated in women during orgasm and AVP in men increase during sexual arousal adding validity to the notion that sexual activity indeed reinforces the bond. In the postpartum state when sexual activity and desire decreases probably as a trade off in reproductive interests. This appears to be mediated by OT through activation of reward centers in the VTA.[35]
Thus, the neuroendocrine system for sexual attraction and partner attachment appear to work in tandem in a monogamous species motivating individuals to prefer a specific mating partner and also motivating them to form an attachment to this mate. In nonmonogamous species, sexual attraction and partner attachment appear to operate independently. The neuroendocrine networks that mediate these complex relationships appear to themselves be complex, flexible, and interdependent and facilitate individuals of myriad species with the range of motivations, emotions, and behaviors necessary to pursue their species-specific reproductive strategy.[8]
Conclusion
Landmark studies in the prairie vole coupled with functional MRI studies have helped us understand the complex interplay of distinct pathways that mediate sexual attraction, romantic love maternal love, and platonic friendships. Further ongoing research is attempting to delineate the biologic basis of complex traits including fidelity, trust, and spirituality. The role of modification of these neurotransmitters particularly OT in the therapy of autism, in trust deficit and in behavioral diseases requires further delineation. For example, OT is generally regarded as a hormone that enhances trust behavior. However, its administration of OT in women who have a forgiving attitude increases the chance that they may punish betrayal.[36] Thus, the response to therapy with OT (and by inference other peptides) appears to be conditional to differences in the individual. These include sex and hormonal status, variations in the OTR, early experiences, epigenetic changes, and neuroplasticity.[37] Large scale trials will further clarify the role of peptide therapy in modulating behavior. And whether successful or not, manipulating love and emotions hoists a slew of ethical red flags. At the time of this writing, however, romantic love retains its mystique continues to tantalize it has revealed some secrets but has manage concealed the vital.
Financial support and sponsorship
Nil.
Conflicts of interest
There are no conflicts of interest.
References
Article №3
oxytocin and love: Myths, metaphors and mysteries
February 2022, 100107
https://doi.org/10.1016/j.cpnec.2021.100107
aKinsey Institute, Indiana University, Bloomington, USA
bDepartment of Psychology, University of Virginia, Charlottesville, USA
Highlights
- oxytocin is a peptide molecule with functions that support a sense of safety, sociality, as well as survival and reproduction.
- oxytocin is associated with social and neuroimmune solutions to chronic stress.
- The related, but more primitive, peptide vasopressin supports more individualistic survival strategies.
- Controversies and myths surround the properties of oxytocin and love.
Abstract
oxytocin is a peptide molecule with a multitude of physiological and behavioral functions. Based on its association with reproduction - including social bonding, sexual behavior, birth and maternal behavior - oxytocin also has been called “the love hormone.” This essay specifically examines association and parallels between oxytocin and love. However, many myths and gaps in knowledge remain concerning both. A few of these are described here and we hypothesize that the potential benefits of both love and oxytocin may be better understood in light of interactions with more ancient systems, including specifically vasopressin and the immune system. Oxytocin is anti-inflammatory and is associated with recently evolved, social solutions to a variety of challenges necessary for mammalian survival and reproduction. The shared functions of oxytocin and love have profound implications for health and longevity, including the prevention and treatment of excess inflammation and related disorders, especially those occurring in early life and during periods of chronic threat or disease.
Keywords
Oxytocin; Vasopressin; Immune system; Social behavior; Stress; Love
1. Overview
Oxytocin was the first peptide molecule to be biochemically identified [1]. The oxytocin molecule has properties and functions essential to mammalian behavior and to understanding human origins [2] which are only now being described [3,4]. Interest in oxytocin has increased exponentially, especially over the last two decades. At present approximately 30,000 research papers listed on Pubmed deal in some way with oxytocin. However, despite this massive effort, the study of oxytocin remains associated with myths and mysteries.
This perspective essay considers a series of questions which are fundamental to understanding oxytocin, but which also have been sources of repeated controversy and misunderstandings: What is oxytocin and why is this molecule difficult to measure? Is there a unique receptor for oxytocin? Is oxytocin a stress hormone? Are the actions of oxytocin sexually-dimorphic? Why is awareness of vasopressin and the immune system essential to understanding oxytocin? What is social behavior and how does oxytocin influence sociality? And of course, why is oxytocin sometimes called “the hormone of love”?
Oxytocin has effects on social connection, a perception of safety, and also immunology and inflammation. Of particular importance in understanding oxytocin are its interactions with the immune system [5], as well as the anti-inflammatory effects of mitochondria [6,7]. Oxytocin regulates and is regulated by glia [8] and the immune system [9]. Furthermore, oxytocin mediates behavioral and immune consequences that are attributed to the microbiota [10,11].
All of these have consequences for health and wellbeing. As described below, recent studies are helping to explain some of the myths and controversies associated with oxytocin. The mechanisms linking love and oxytocin to each other remain both metaphorical and mysterious, but a mystery worth exploring.
2. So, what exactly is oxytocin?
The “what is oxytocin” question should be easy to answer. The use of the word oxytocin is not new. The website of theNational Institutes of Mental Health states that the term oxytocin was already in the use in the 1800s and perhaps as early as the 1500s.
https://www.nimh.nih.gov/research/research-conducted-at-nimh/research-areas/clinics-and-labs/lcmr/snge/vpot/some-selected-history-of-oxytocin-and-vasopressin. As originally used, the word oxytocin appears to have referred to a process of “swift birth.” However, in current usage oxytocin most often describes a specific molecule.
Although modern dictionaries usually translate the derivation of the word oxytocin as “swift birth,” as described in an email from Roger Guillemin, “oxytocin”, the English spelling, may have been a “mistaken translation”? (Box 1). Dr. Guillemin is a Nobel laurate and sometimes described as father of the field of neuroendocrinology. Guillemin suggested that oxytocin translates to “sharp” birth and proposed that “ocytocin” (still used in French) would have been a more accurate translation of the Greek symbols intended to describe the functional capacity of this molecule to induce a quick birth.
BOX 1
THE MYSTERIOUS ORIGINS OF THE WORD “OXYTOCIN. (An historical aside)
This excerpt is from an email correspondence with Roger Guillemin. It came following a talk I had given at the Salk Institute. In that presentation I showed a portrait of Antoine Lavoisier(father of modern chemistry), his wife, Marie-Anne, and instruments for collecting oxygen – a picture I sometimes used to symbolize the notion that loving relationships are based on biochemistry.
…. As per Lavoisier, the word oxygen comes from the Greek ὀξύς/oxys, acid, sharp and γενής/genês, generates, because Lavoisier thought that the substance oxygen was involved in producing acids in combination with other substances (still true though not exclusive …). Oxytocin is a totally different story. It is actually a mistake for ocytocin.
The etymology is ὠκύς/ocys fast and τόκος/birth, delivery. Note the κ (kappa) always translates as c (ocytocin) different from the ξ (ksi) always translated as x as in oxygen. I went over on and on with texts in Greek, Herodotes, Hippocrates, where I found the words ὠκύς τόκος, as in 'ωκυτοκιος ′ωκυτοκιοv ὠκυτόκου all in phrases referring to speedy birth, never as ὀξύς as for oxygen (acid generating) So the correct word is OCYTOCIN.
Well maybe here's the last word in a story told me, many years ago by Roger Acher whose name you recognize of course... At some meeting duVigneaud was talking about his early work on isolation of oxytocin when this French participant (?) got up and said …
“ Sir, with all due respect please note that the correct word is ocytocin and I can show the etymology” to which duVigneaud is supposed to have answered. “ listen I don't care about all your grammar... all I know is that stuff comes from the pituitary of oxen... so that's oxytocin by me..." I knew duVigneaud quite well and I bet the story is true …
I still have not been able to find out who first originated OXYTOCIN.
—Roger Guillemin May 29, 2013.
Genomic studies, tracing the evolutionary origins of these molecules, further question the accuracy of this name [12]. Theofanopoulou and her colleagues have suggested the need for a “universal nomenclature for oxytocin-vasotocin ligand and receptor families”. One family, that they propose to call “vasotocin,”, would consist of the molecules that are currently called vasotocin and vasopressin (Table 1). A second chemical family they propose to call “oxytocin”, would consist of oxytocin, mesotocin and isotocin. These authors argue that a more accurate naming system would be based on the relative affinity of ligands to receptors. Following this system, the molecules now called mesotocin and isotocin, would be called oxytocin. Of course, calling these three structurally different molecules “oxytocin”, offers opportunity for other sources of confusion and controversy.
| Amino acid position | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | Expressed in (among others)a |
|---|---|---|---|---|---|---|---|---|---|---|
| Vasotocin | Cys- | Tyr- | Ile- | Gln- | Asn- | Cys- | Pro- | Arg- | Gly (NH2) | Non-mammalian vertebrates and fetuses |
| Vasopressin | Cys- | Tyr- | Phe- | Gln- | Asn- | Cys- | Pro- | Arg- | Gly (NH2) | Mammals |
| Oxytocin | Cys- | Tyr- | Ile- | Gln- | Asn- | Cys- | Pro- | Leu- | Gly (NH2) | Mammals |
| Mesotocin | Cys- | Tyr- | Ile- | Gln- | Asn- | Cys- | Pro- | Ile- | Gly (NH2) | Non-eutherian tetrapods & birds |
| Isotocin | Cys- | Tyr- | Ile- | Ser- | Asn- | Cys- | Pro- | Ile- | Gly (NH2) | Ray-finned fishes |
|
|
Oxytocin and its targets have consequences that remain essential to coordinating mammalian behavior and reproduction with environmental demands, including functions such as defense in the face of pathogens [13], immunological responses, mitochondrial functions and basic cellular metabolism [6]. Oxytocin also can function as a neurotransmitter and neuromodulator [14]. Perhaps even calling oxytocin a “hormone” is a misnomer?
3. The variable chemistry of the molecule called oxytocin
In 1953 oxytocin and vasopressin, were identified biochemically [1]. The gene for oxytocin receptor have been well understood for several decades. But again, the story is not that simple.
Canonical forms of oxytocin and vasopressin consist of peptide molecules composed of nine amino acids, with a three amino acid tail (implicated in ligand binding to the receptor) and a six amino acid ring (implicated in coupling with G-coupled protein receptors (GCPR) [17]. The ring in this structure is formed by the disulfide bonds in two cysteines (positions 1 and 6). (Fig. 1; Table 1). Vasopressin differs from oxytocin by two amino acids, one at position 3 (in the ring) and one at position 8 (in the tail). Vasotocin, considered the ancestral peptide from which both evolved, differs from vasopressin by one amino acid at position 3 and from oxytocin by one amino acid at position 8.
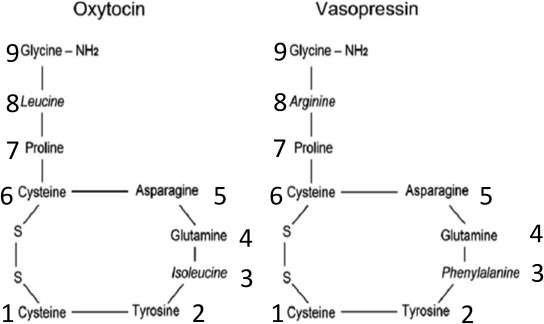
One interesting difference between the structure of oxytocin versus other major peptides in this family (Table 1) is the unique sequence of three amino acids in the tail at positions 7–9 (PLG: proline-leucine-glycine), which is also known as melanocyte-inhibiting factor (MIF-1). MIF-1 has pain-modulating effects via interactions with opioid and dopaminergic systems (reviewed [3]). The structure of MIF-1 is not found in vasopressin or mesotocin. This may be just one of many clues regarding what makes oxytocin unique.
A common feature shared by canonical oxytocin and other members of this chemical family is the presence of the disulfide bonds connecting the two cysteines and creating the ring structure of these molecules. The rings in these peptides are capable of opening to create linear molecules. It has been proposed that the linear form of oxytocin or fragments of oxytocin may be of particular importance to its anti-inflammatory and anti-oxidant effects [18].
Oxytocin with leucine (at position 3) and isoleucine (at position 8) is found in most, but not all, mammals. However, species-specific variants in the amino acid sequences for oxytocin have been discovered recently and associated with different patterns of sociality. For example, amino acid substitutions at position 8 (and in some cases also position 3) exist in New World versus Old World monkeys. Substitutions of one or more amino acid also may produce not only different binding affinities, but also differential capacities to desensitize the oxytocin receptor, and thus potentially allow longer durations or strengths of action.
It has been suggested that in comparison to canonical oxytocin, the oxytocin-like variants found in highly social New World monkeys may be not only better able to bind to the oxytocin receptor, but also less likely to bind to the vasopressin V1a receptor [19]. Associated with these novel variants is male parental behavior, a defining behavioral trait of many species of New World monkeys. Remarkably, after receiving one novel form of oxytocin, described as Pro8-oxytocin and previously associated with parental behavior and social monogamy in New World monkeys, nonparental male rats were more likely to show parenting behavior toward infants [20]. Findings of this sort continue to support the notion that, irrespective of nomenclature, the exact chemistry of oxytocin-like molecules matters and small structural differences can have functional consequences.
4. Are measurements of oxytocin meaningful?
The structural characteristics and especially the “sticky” sulfur bonds in oxytocin influence its functional availability within tissues. The capacity of oxytocin to bind to other biological entities also has contributed to controversy regarding the quantification of oxytocin. Current antibody-based methodologies provide snapshots of hormone levels, and are most often used in plasma or saliva. These have proven useful when considering correlations with behavioral patterns and responses to external stimuli. For example, levels of oxytocin and vasopressin rise after cold-stressors or music [21] and oxytocin levels have been individually correlated with electrical activity in the brain specifically in response to social cues [22]. These measurements are of most value within a given study [23]. Furthermore, as described below other molecules capable of transporting oxytocin, including RAGE (“receptor for advanced glycation end-products”), may create biochemical interference in antibody-based assays [24,25]. This in turn is likely to contribute to variation in estimates of the amount of endogenous oxytocin in bodily fluids.
Perhaps, rather than dealing with the dynamic and variable characteristics of the oxytocin system as controversies or myths [26], it may be more helpful to conceptualize bodily fluids as dynamic delivery and/or storage systems. These in turn function as components of the body's active defense systems, providing local availability of oxytocin, especially in response to threatening conditions such as infection or inflammation [27].
5. Is oxytocin a “pituitary” hormone?
Sir Henry Dale identified the presence of a substance in pituitary extracts that could facilitate uterine contraction. Dale is usually given credit for discovering oxytocin, although in his original 1906 paper he did not specifically call this molecule “oxytocin” [28]. Oxytocin does have classical hormonal functions, since it is stored in the posterior pituitary, released into the blood stream, and carried to target tissues throughout the body. However, oxytocin is not actually synthesized in the pituitary gland, but in fact is produced in the hypothalamus.
Hypothalamic neurons which synthesize oxytocin and vasopressin do extend processes to the posterior pituitary gland; there these peptides are released into the systemic circulation, coordinating birth, lactation, sexual responses, water and mineral homeostasis, cardiovascular function and metabolism in bones and muscles, to name only a few. Hypothalamic cells also release both oxytocin and vasopressin directly into the brain with local effects on specific tissues and functions [29]. For these reasons oxytocin is now commonly described as a “hypothalamic neuropeptide”, rather than a pituitary hormone. But perhaps even that distinction is not totally accurate?
6. Is oxytocin primarily a hypothalamic hormone?
Although the hypothalamus is often cited as the primary source of oxytocin, the oxytocin molecule is synthesized in other parts of the body. In fact, based on gene expression for oxytocin, in the female rat uterus near the time of parturition oxytocin levels are reportedly 70 fold higher than any other place in the body, including the hypothalamus. As originally explained by Lefebvre, Zingg and their colleagues,“endogenous circulating OT appears not to participate in the induction of labor… and the finding of OT messenger RNA and peptide in the uterus suggests a solution for this paradox. During parturition, OT may act primarily as a local mediator and not as a circulating hormone”[30].
Oxytocin also appears in extremely high levels at sites of inflammation. In ovarian cancer tumors the levels of oxytocin were up to 200 times greater than in plasma [27]. Oxytocin levels measured in these tumors were negatively associated with an inflammatory cytokine, interleukin-6 (IL-6). A higher level of oxytocin within tumors also was correlated with mortality in advanced cases of ovarian cancer, an effect that was mediated by IL-6. This research further implicates locally synthesized oxytocin as an anti-inflammatory agent and an important component of the immune response [31].
7. Is there a “unique” oxytocin receptor and why does this matter?
In a comprehensive review of the oxytocin receptor Jurek and Neumann [32] state… “The many facets of oxytocinare, on a molecular basis, brought about by a single receptor.” When compared to related peptides, oxytocin generally binds most readily to the oxytocin receptor and GPCRs are an important mechanism for oxytocin's action. GPCRs have the capacity to respond to different ligands and transduce information across cell membranes, triggering subcellular processes [4] and thus influencing downstream transcription. Differences in subcellular signaling can have an array of functions that may include the expression of social behavior [33].
A separate set of GCPRs are described as vasopressin receptors. Of these the vasopressin V1a and V1b receptors found in the nervous system, can be targets for both oxytocin and vasopressin [34,35]. In the genetic absence of oxytocin the vasopressin system may assume some, but not all, of the functions of oxytocin, acting in part via sensitized vasopressin receptors [36]. As described below interactions between oxytocin and vasopressin are most obvious under threatening conditions.
Receptors responsive to oxytocin are abundant in the nervous, reproductive and immune systems including on microglia and astrocytes [9]. These receptors are cell-type specific [14]. Awareness of local effects of oxytocin is providing critical information regarding tissue-specific actions of oxytocin, especially in the nervous system [4]. In one of many recent examples, oxytocin receptors on astrocytes in the amygdala have been implicated in a neural circuit that can reduce anxiety [37]. Studies such as these provide additional evidence for interactions between oxytocin, components of the immune system and neural function.
The crystal structure of the GCPR oxytocin receptor was recently described [38], offering fresh clues to the specific capacity of different kinds of molecules to bind to the oxytocin receptor and moderate its function. This research revealed that the functions of the oxytocin receptor are affected via positive allosteric stimulation by magnesium (Mg++) and other divalent cations, as well as negative allosteric regulation by sodium (Na+).
Some of the short-term consequences of oxytocin act via ion channel receptors [39]. Gaseous molecules, including hydrogen sulfide (H2S) and nitric oxide (NO), also have been implicated in the rapid actions of oxytocin, helping to explain the role of oxytocin in the regulation of both the autonomic and cardiovascular systems [40]. Clearly, GPCRs - even those originally described as oxytocin receptors - are only one mechanism through which oxytocin functions. The availability of multiple mechanisms for responding to oxytocin, often with different time courses, has consequences for the effects of oxytocin throughout the body.
8. Is oxytocin a “female” hormone?
The first myth concerning oxytocin's functions arose when the molecule was identified as a female reproductive hormone, without known effects in males. Male and female mammals do use different biological and behavioral strategies and functions both for reproduction and for survival. Patterns of sex differences are reported in the production of oxytocin, the oxytocin receptor and their functional consequences [41]. Dozens of studies also reveal sex differences in response to exogenous peptides, including oxytocin and vasopressin [42,43]. These sex differences are managed in part by steroid-peptide interactions [44,45], and also may reflect sex differences in the functions of mitochondria and the immune system [8,46], in turn relevant to managing inflammation and reproductive demands, as well as sexual differentiation [47].
Sex differences in oxytocin and vasopressin are especially likely to be detected in response to stressful experiences during the perinatal period [48]. Under these conditions vasopressin has been more often implicated in males [49,50], while CRF and adrenergic systems (including norepinephrine) may be critical to managing chronic stressors in females [51]. However, a role for oxytocin in coping with stress also has been identified in male rodents [53]. This involves the capacity of estrogens and androgenic metabolites (specifically 3 β diol) to differentially stimulate estrogen receptors, including the estrogen receptor (ER) β. The ER β found in the hypothalamus can affect the release of oxytocin, and reduce measures of anxiety and inflammation. Thus, although usually identified with females, steroidal systems including estrogen receptors and oxytocin have important consequences for stress management in males.
Sex differences in physiology and behavior also appear during reactions to stressful experience during the perinatal period and there is increasing evidence implicating inflammation in sexual differentiation [47]. Given the importance of oxytocin as an anti-inflammatory molecule, it is likely that oxytocin moderates certain aspects of masculinization, possibly reducing the neural consequences of androgens. Differential roles for sex steroids and the oxytocin-vasopressin system also can have major consequences for species differences in social behavior [44].
Complex interactions among genetic sex, gonadal steroids, stress and immune factors and lifespan development are difficult to parse experimentally. However, these are very important pathways to consider since they help to sculpt the connections among social support and wellbeing.
9. Is oxytocin an “anti-stress” hormone?
Oxytocin has been described as a stress hormone [53], a stress-coping molecule [54], or both [29]. In general, oxytocin has central roles in coping with challenge. Oxytocin modulates stress reactivity and facilitates restoration following periods of challenge and has many interactions with the hypothalamic-pituitary-adrenal (HPA) axis, immune and autonomic systems [3,32]. In addition, time is a critical factor in understanding this aspect of oxytocin's functions. Oxytocin and vasopressin are both released under conditions of acute demands, including sexual orgasm [54], ejaculation [56], birth [57], pair bond formation [58], intense exercise [59], severe pain or shock-trauma [40,60] and sodium challenge [61]. In all of these cases at least one of the roles for oxytocin and social connections may be to calm the organism and reduce inflammation [62], helping systems return to homeostasis, while also predicting upcoming allostatic demands [63].
Oxytocin also has the capacity to increase its own synthesis and release [64]; this feature of oxytocin accounts in part for its extensive use to facilitate parturition [57]. Even in nonpregnant animals, stimulating the release of oxytocin or administering exogenous oxytocin is capable of increasing the subsequent synthesis of endogenous oxytocin [65]. The ability of oxytocin to feed forward and increase its own synthesis could be critical to coping with chronic challenges - especially in a social context. Vasopressin also has the capacity to feed forward, an effect that can be increased by androgens [66]. However, in general the long-term effects of vasopressin are different, and in many contexts opposite from those of oxytocin (reviewed [32]).
The effects of oxytocin during responses to challenge occur against a functional background of interactions between vasopressin and corticotropin releasing hormone (CRH). Vasopressin and CRH are often co-localized and can initially amplify the effects of each other, increasing activity in the HPA axis [49]. However, during chronic stress vasopressin, versus CRH, provides a mechanism allowing “escape” from glucocorticoid-regulated negative feedback [50]. Under chronic stress, oxytocin's capacity to downregulate defense systems may be especially relevant, allowing social and psychological safety to modulate both the HPA axis and emotional reactivity.
Particularly important to understanding the stress-related consequences of oxytocin is its capacity to regulate chronic pain [67]. Whether oxytocin's role in reducing pain is via the oxytocin and/or vasopressin receptor is still being debated [68,[69]. There is evidence that oxytocin can induce analgesia through effects on the vasopressin V1a receptor [70]. Vasopressin may have a different time course of action, serving in the early stages of a stressful experience when a pro-inflammatory response is important. Oxytocin seems to be particularly relevant during repeated or chronic stressors, possibly through its capacity to counteract the pro-inflammatory effects of vasopressin as well as the capacity to signal social safety [8].
Positive or negative experiences can adjust reactivity in the HPA axis, as well as the oxytocin system across the lifespan. This is especially apparent around the time of birth, when oxytocin can upregulate the oxytocin receptor [71]. For example, postnatal exposure to oxytocin or parental nurture interferes with de novo methylation of the oxytocin receptor which occurs in the postpartum period. Under these conditions oxytocin and/or positive nurture is associated with upregulation of the expression of the oxytocin receptor and increasing indications of sociality in later life [72,73]. The developmental effects of vasopressin are less well studied [74]. However, research in rodents suggests that - especially in males - early exposure to vasopressin upregulates emotional reactivity and aggression, with consequences across the life cycle [75,76].
Both immediate demands and the history of each individual also can influence responses to oxytocin and vasopressin. For example, research, primarily in nonhuman animals suggests that vasopressin's effects are most obvious after a history of adversity, especially when compared to high levels of early nurture [77]. The social history of an individual can alter the threshold for responding to oxytocin, vasopressin and other stress-related molecules [74]. This likely reflects adjustments to the sensitivity of both vasopressin and oxytocin systems, with later consequences for the capacity to manage inflammation and oxidative stress, of importance in many stressful contexts.
10. Ancient molecules and the evolution of a social solution to the stress of life
Many of the properties of oxytocin are best appreciated in the context of its' evolutionary history and interactions with even more ancient molecules. oxytocin-like molecules facilitate social and reproductive interactions in both vertebrates and invertebrates [78]. Precursors for oxytocin and the related peptide vasopressin existed prior to the preCambrian explosion when atmospheric oxygen levels on Earth increased, permitting a dramatic increase in multicellular organisms. The evolution of these peptides was associated with the transition to living on dry land, the need to regulate bodily fluids and nutrients and to deal with exposure to elevated levels of oxygen [79]. However, oxygen is potentially dangerous. oxytocin's anti-inflammatory and anti-oxidant properties also may have helped to protect the mammalian nervous system during exposure to atmospheric oxygen.
Oxytocin evolved from mesotocin possibly as long as 400 million years ago [81], preceding and facilitating the origins of mammals and lactation [82]. Many types of organisms exhibit viviparity, but milk from a mammary gland and the genes responsible for milk production are uniquely expressed in Mammalia, including monotremes, marsupials and eutherian mammals [83]. Supporting a gradual evolutionary change, monotremes, egg-laying mammals that also lactate, produce both mesotocin and oxytocin.
Peptides structurally identified as canonical oxytocin have been identified in jawless fish and in cartilaginous dogfish [82]. However, the oxytocin found in fish does not appear to have been the ancestral source of mammalian oxytocin. It is hypothesized that oxytocin in fish arose as a function of “random genetic drift,” while “selection evolution” led to the independent emergence and maintenance of oxytocin in mammals [80].
Many other biologically active molecules, including common neurotransmitters and variations on the CRH molecule existed prior to the preCambrian explosion [84]. In that context the evolution of the specific molecule known as oxytocin is comparatively recent. Furthermore, in comparison to the “primitive” anatomy of vasopressin-secreting neurons, hypothalamic neurons producing oxytocin have very complex branching patterns [79], forming feedback networks reaching from the brainstem to the cortex [85].
Oxytocin's functions also are intertangled at many levels with the ancient immune system, helping to manage challenges across the lifespan [5,86]. For example, RAGE is a component of the immune system that serves as a carrier molecule facilitating oxytocin's movement across tissues, including placental and blood brain barriers [87]. CD 38 is another immune system molecule that regulates oxytocin. CD 38 is located on cells throughout the immune system and was initially described as an “immune cell marker”. It is now understood that CD 38 also is essential for the calcium-dependent release of oxytocin. Both CD 38 and RAGE also are implicated in maternal behavior and other forms of sociality [5]. Research on these interactions supports the importance of interactive functions of oxytocin, the immune system and positive sociality. Although CD 38 is essential to the normal functioning of oxytocin system, apparently CD 38 is not required by the vasopressin system [86], suggesting another unique feature linking the oxytocin and immune systems.
11. The effects of oxytocin and vasopressin are hierarchical and context-dependent
In healthy individuals who have experienced high levels of early nurture, oxytocin can have immediate benefits in response to stressors, including allowing a return to homeostasis [33], more accurate appraisals of threat versus safety and future risks [88] and prediction of future allostatic demands [63]. However, under extreme or repeated stress, and especially in individuals that have experienced high levels of adversity in early life, the consequences of high levels of oxytocin are less predictable. In part this could be because under these conditions vasopressin receptors are triggered [74,89]. Interactions among oxytocin and vasopressin probably help to explain what have been called “paradoxical” effects of oxytocin [90], including negative responses to exogenous oxytocin [35].
During social encounters oxytocin potentially inhibits the defensive and more primitive actions of vasopressin and other stress-related pathways. Oxytocin, may reduce the perception of threat, allowing animals to engage in prosocial interactions and in some cases develop selective relationships. However, under conditions of adversity or trauma across the lifespan, pathways dependent on vasopressin may dominate, even when oxytocin is administered or otherwise abundant. For example, during a difficult childbirth, maternal physiological systems may default to mechanisms that are regulated by vasopressin [91] and/or effects of oxytocin on the vasopressin receptor [69,70]. In another example, after receiving exogenous oxytocin individuals with a history of early life adversity or mood disorders may show negative emotional reactions, resembling those that might be expected following vasopressin [92,93].
In summary, oxytocin both regulates and is regulated by components of the immune system including glia [89]. Evidence is growing for other relationships between oxytocin and the immune system [5] as well as the capacity to affect the anti-inflammatory actions of mitochondria [67]. Oxytocin also has consequences for the release of gaseous transmitters, such as H2S or NO, with potential protective effects throughout the body [40]. Furthermore, oxytocin apparently mediates behavioral and immune consequences that are attributed to the microbiota [10,11].
12. What does it mean to be called a “social neuropeptide”?
It has become common to described oxytocin as a social neuropeptide and hundreds of studies describe the role of oxytocin and vasopressin in the neurobiology of social behavior. However, exactly what this means needs additional consideration. States of perceived threat, versus safety, influence the capacity for and expression of positive social behaviors (reviewed [89]). Constructs like social behavior [94], social recognition [95], social salience [96] and even maternal behavior [97] are difficult to differentiate from the general tendency of mammals to avoid danger and seek safety, especially in response to novel social stimuli.
Awareness of emotional context and the history of each individual, including prior experiences of safety versus threat, are particularly critical in understanding the consequences of oxytocin and vasopressin [98]. However, the capacity of oxytocin to influence responses to social cues may not be readily apparent until the individual is faced with a challenge, including for example a newborn infant. As another example, cues of sickness can be powerful in eliciting social avoidance [99] [8]. Oxytocin can override social avoidance [100] promoting social interactions even in the face of biological threat [13]. Here again, cross-reactivity among oxytocin, vasopressin and the immune system offers mechanisms for adaptive responses to specific social stimuli [101].
13. Is oxytocin “the hormone of love?”
Single molecules – even ones as versatile as oxytocin - cannot explain the intricacies of a behavioral construct like love. A host of neural systems, neurotransmitters and neuromodulators, especially catecholamines, indoleamines, acetylcholine, endogenous opioids, sex steroids, molecules of the HPA axis, including CRF and corticosterone, and inflammatory cytokines, interact in the regulation of the oxytocin system [102]. For example, of particular importance are GABA [103], serotonin [104], dopamine [105], and opioids [106,107]. All of these chemicals are more ancient than oxytocin and vasopressin [84]. However, most of these molecules and their evolved interactions with mammalian biochemistry have been implicated in the neurobiology of love [108], as well as physiological reactions to threat.
Oxytocin does support physically intimate forms of sociality and nurture and plays a critical role in infant feeding. Unique properties of oxytocin also were involved in the evolution of the capacity to form lasting attachments [109]. Acting on various target tissues and neuroendocrine pathways oxytocin helps to regulate emotional states including those that are experienced by the human nervous system as love. In this context, oxytocin became known as “the hormone of love” [110].
Mammalian maternal behavior, acting through oxytocin, was originally suggested as a model for “love” [111]. However, studies of genetically mutant mice unexpectedly revealed that maternal care continued in the absence of oxytocin and its receptor [112]. Parental behaviors are the product of a complex and potentially redundant neurobiology. In a strict sense oxytocin is not “the” hormone of mother's love [113,144].
Young mammals are especially salient stimuli, with the potential to attract attention and elicit care, apparently even in the absence of oxytocin. In some contexts, young animals, including offspring, also can serve as threats, inducing avoidance or attack. To engage in parenting, mammals must selectively overcome cues of threat, while permitting defense against possible dangers. For example, early research showed that responses to infants increased in anosmic virgin female rats, suggesting the olfactory cues from pups inhibited maternal-like behaviors [115].
However, oxytocin may serve mammals as a kind of insurance policy against over reaction to their young or other environmental stressors [116]. For example, a postpartum surge in oxytocin helps to expel the placenta and could support initial bonding between the mother and child [117]. Whether the mechanisms through which oxytocin serves to influence approach behavior are specific to parental-young interactions, or perhaps represent a more general reduction in reactivity to threats, deserves additional research [118,119].
14. Is oxytocin a metaphor for love and safety?
The word “love” has many meanings and interpretations [120,121]. Love can be described as a metaphor, albeit with physical and psychological manifestations [122,123]. Of course, even repeated associations across different levels of analysis do not prove causation. However, within science most definitions of love involve selective behaviors and attachments [109]. Using this limited definition, remarkable parallels can be identified between the functions and properties of oxytocin and love (Table 2). The features and functions of oxytocin and social attachments suggest that these apparently diverse constructs have shared roots. As described here, it can be argued that oxytocin is a component of an embodied biological system that supports the benefits of secure relationships.
| FUNCTIONS (among many) | LOVE | OXYTOCIN |
|---|---|---|
| MODERN (evolutionarily recent) | + | + |
| Associated with SELECTIVE sociality & bonds | + | + |
| Supporting parental investment | + | + |
| Metaphor for SAFETY | + | + |
| Selectively rewarding | + | + |
| Anti-inflammatory/Anti-oxidant | + | + |
| Anxiolytic/Analgesic | + | + |
| Allows immobilization without fear | + | + |
| Sexually dimorphic | + | + |
| Epigenetically tuned and Context dependent | + | + |
Both oxytocin and love are linked to the health benefits of safety including emotional and physical support [124]. Both also have adaptive interactions with more primitive and individualistic systems, including processes needed to manage threat, inflammation and disease. These more archaic mechanisms support individual survival in the short-term, but prolonged activation of the immune system can be dangerous for long-term health.
Oxytocin and vasopressin interact with other defensive systems, including the autonomic and immune systems, and also regulate the emotional states necessary for perceptions of safety [[63],[63]]. These coordinate the capacity of an organism to selectively exhibit behavioral and emotional states necessary for active versus passive defense, including the alertness and mobility needed for escape from threat, danger or pain [126]. Peptide-mediated behavioral and autonomic functions also can influence the capacity for the voluntary immobility without fear, required for social interactions including maternal and sexual behaviors [127].
Of particular importance to mammalian survival are both early nurture and lasting and secure relationships [128] which may be facilitated by oxytocin [129]. Oxytocin has an essential role in lactation, facilitates the capacity to give birth and may allow mammals to become attached to their infants even under conditions of extreme challenge, such as those associated with human birth and child rearing [116]. Lactating women are uniquely able to manage stressors [130]. A postpartum surge in oxytocin helps to expel the placenta and could support initial bonding between the mother and child [117]. The development of offspring also is modulated by oxytocin [131], with effects that can be protective after a surgical birth [132]. Furthermore, infants reared on human milk receive a cocktail containing optimal nutrition, as well as oxytocin itself and probiotic bacteria [7,11].
The oxytocin system is an intrinsic component in the capacity of experiences, including both nurture and adversity, to manage adaptations in the mammalian body. The epigenetic consequences associated with modifying the oxytocin system can be detected across the lifespan, but especially around the time of birth when newborns must anticipate the demands of their future environments [7,103]. Early life is a period when mammals are particularly sensitive to the need for nurture and when the effects of oxytocin are most likely to create epigenetic change [[71], [72], [73]].
Oxytocin and positive social experiences play a critical role in brain development [94,133]. Early experiences also are implicated in the capacity to create lasting attachments [134]. As discussed elsewhere, oxytocin was permissive in the evolution of the human nervous system [2], with the resultant large neocortex necessary for consciousness, language and even spirituality. This capacity in modern humans is required for relationships to be experienced and expressed [135]. Furthermore, accurate appraisal of future threats [136], the relevance of social stimuli [119], and the development of a complex brain and autonomic nervous system demand oxytocin [2].
Both the health benefits of oxytocin and the capacity for love rely on interactions with more ancient physiological systems, including the immune and vasopressin systems [3,81,137]. The patterns of behavior regulated by vasopressin also are adaptive and in the functional absence of oxytocin the effects of vasopressin may be associated with fear, self-defense and in some cases aggression [89]. Understanding the factors that regulate these reactions provides novel insight into the complexity, power and possible dangers associated with love or its absence.
Biologically-active molecules, such as oxytocin, are sometimes treated as if they are manifestations of a well-understood chemical reality. Of course, oxytocin is part of a larger physiological equation. Here we have examined specific examples of interactions of oxytocin with vasopressin and the immune system. These have implications for all aspects of human health, and especially therapeutic strategies for the prevention and treatment of diseases associated with stress, adversity, trauma and inflammation.
15. Gaps in knowledge and a work in progress
Oxytocin and love have shared properties and functions (Table 2). Accurately calibrating reactions in response to or in anticipation of challenge are of particular importance to survival and eventually reproduction 138. A general feature of both oxytocin and love is the capacity to modulate more primitive defense systems including threat or fear. Many of the biological associations between oxytocin and love are only apparent under conditions of physiological challenge. Loving and secure relationships, in part through the dynamic actions of oxytocin, are especially critical in the face of threat [89]. The effects of both oxytocin and secure relationships may be most easily detected in response to stressful and chronic experiences, including sickness [13,99] and social isolation [8]. Oxytocin also may help to explain the benefits of companion animals [139], including the processes that led to canid domestication [140].
Oxytocin's role in reducing emotional threat may be of particular importance in understanding the consequences of social isolation or during disrupted relationships, and especially in women [42]. Animal research also indicates that the mechanisms through which isolation is so devastating involve dynamic and sexually-dimorphic changes in oxytocin [65,141,142]. Interactions between oxytocin, vasopressin and immune systems are important to complex “paradoxical” effects of exogenous oxytocin [89], and could help to explain why it has proven difficult to create pharmaceuticals based on these peptides [3]. However, the nature of these interactions remains at present poorly understood.
Discussed elsewhere is evidence for a major role of the autonomic nervous system in mediating social connection and emotion regulation [3,125.]. The autonomic system also regulates immunity and is implicated in autoimmune disorders [143]. Oxytocin and vasopressin both affect the complex coupling of sympathetic and parasympathetic functions [63,144.]. The relationship between oxytocin, vasopressin and the autonomic nervous system deserves deep analysis.
Excess inflammation and oxidative stress have been implicated in virtually every known physical and emotional disorder [145]. Oxytocin functions in part through its capacity to serve as a component of mammalian defense systems with both anti-inflammatory and anti-oxidant effects [146] [147]. The detrimental effect of oxygen also is managed in part by the anti-inflammatory and anti-oxidant properties of oxytocin. This may be a clue to an ancient and reoccurring link between oxytocin and oxygen. This link is found in responses to immune challenges and excess inflammation, including mitochondrial activity [6,148], responses to pathogenic viruses [149], and in protection of the fetus during live birth [103].
Broad metaphorical meanings have been attached to both the concept of love and the functions of oxytocin. Compared to other biologically active molecules oxytocin is recently evolved. Oxytocin links individuals to others, and also supports the benefits of sociality. Survival strategies based on connection, cooperation, the support of offspring and perceived safety are critical to emotional and physical health [150]. There is evidence that the mechanisms for the benefits of oxytocin and secure relationships offer potential, but at present largely untapped, insights into disease, cellular aging and longevity [151,152].
Myths and mysteries surround love. Despite popular assumptions that “love overcomes fear” and “love heals,” as a subject for serious study love may be considered frivolous. We argue here that oxytocin has broad consequences that resemble in part an embodied metaphor for love. However, like love, the oxytocin system also has proven difficult to identify, measure and study and the controversies associated with oxytocin are far from resolved. The same adaptive biochemical properties that make oxytocin and love difficult to study may also help to explain their potential as components of “Nature's medicine” [3].
A deeper awareness of the biology of relationships is essential to understanding what it means to be “human.” Mechanisms through which these systems are integrated has profound implications for human health [3]. However, interactions among the pieces of the peptide-immune-behavioral puzzle are only now being recognized. Embedded in the biology of love and oxytocin are secrets to the social solutions that optimize living in a dangerous world. Put simply, without oxytocin and love mammals may survive, but not thrive. How this occurs remains one of life's great mysteries.
Declaration of interests
None.
Acknowledgments
Research described here from the author's laboratory was funded by the National Institutes of Health, most recently by HD P01 07575 and HD R01 098117 and support from the Fetzer Institute. The editorial suggestions and other supportive inputs of Stephen Porges, Alex Horn, John M. Davis, Marcy Kingsbury and Robert Dantzer are gratefully acknowledged. Because of space limitations I apologize to investigators whose primary work is not fully recognized and refer the reader to additional references in earlier reviews.
References
The sequence of amino acids in oxytocin, with a proposal for the structure of oxytocin
J. Biol. Chem., 205 (2) (1953), pp. 949-957
Google Scholar
Oxytocin pathways and the evolution of human behavior
Annu. Rev. Psychol., 65 (2014), pp. 17-39, 10.1146/annurev-psych-010213-115110
Google Scholar
Is oxytocin “Nature's medicine”?
Pharmacol. Rev., 72 (4) (2020), pp. 829-861, 10.1124/pr.120.019398
Google Scholar
The multiple faces of the oxytocin and vasopressin systems in the brain
J. Neuroendocrinol. (June 17, 2021), Article e13004, 10.1111/jne.13004
Google Scholar
CD38, CD157, and RAGE as molecular determinants for social behavior
Cells, 9 (1) (2019), p. E62, 10.3390/cells9010062
Google Scholar
Mitochondria, oxytocin, and vasopressin: unfolding the inflammatory protein response
Neurotox. Res., 36 (2) (2019), pp. 239-256, 10.1007/s12640-018-9962-7
Google Scholar
The inflammatory event of birth: how oxytocin signaling may guide the development of the brain and gastrointestinal system
Front. Neuroendocrinol., 55 (2019), p. 100794, 10.1016/j.yfrne.2019.100794
Journal Article
Google Scholar
Sickness and the social brain: love in the time of COVID
Front. Psychiatr., 12 (2021), p. 633664, 10.3389/fpsyt.2021.633664
Google Scholar
Oxytocin, dopamine, and opioid interactions underlying pair bonding: highlighting a potential role for microglia
Endocrinology, 162 (2) (2021), 10.1210/endocr/bqaa223
Google Scholar
The role of the microbiome in the neurobiology of social behaviour
Biol. Rev. Camb. Phil. Soc., 95 (5) (2020), pp. 1131-1166, 10.1111/brv.12603
Google Scholar
Oxytocin and the microbiome
Curr Opin Endocr Metab Res, 19 (2021), pp. 8-14, 10.1016/j.coemr.2021.04.006
Google Scholar
Universal nomenclature for oxytocin-vasotocin ligand and receptor families
Nature, 592 (7856) (2021), pp. 747-755, 10.1124/pr.120.019398
Google Scholar
Pathogens, odors, and disgust in rodents
Neurosci. Biobehav. Rev., 119 (2020), pp. 281-293, 10.1016/j.neubiorev.2020.09.037
Google Scholar
Molecular basis of oxytocin receptor signalling in the brain: what we know and what we need to know
Curr Top Behav Neurosci, 35 (2018), pp. 3-29, 10.1007/7854_2017_6
Google Scholar
Sequence and organization of coelacanth neurohypophysial hormone genes: evolutionary history of the vertebrate neurohypophysial hormone gene locus
BMC Evol. Biol., 8 (2008), p. 93, 10.1186/1471-2148-8-93
Google Scholar
Oxytocin: the great facilitator of life
Prog. Neurobiol., 88 (2) (2009), pp. 127-151, 10.1016/j.pneurobio.2009.04.001
Google Scholar
Comparison of the pharmacological profiles of arginine vasopressin and oxytocin analogs at marmoset, macaque, and human vasopressin 1a receptor
Biomed. Pharmacother., 126 (2020), p. 110060, 10.1016/j.biopha.2020.110060
Google Scholar
Oxytocin is a principal hormone that exerts part of its effects by active fragments
Med. Hypotheses, 133 (2019), p. 109394, 10.1016/j.mehy.2019.109394
Google Scholar
Leu8 and Pro8 oxytocin agonism differs across human, macaque, and marmoset vasopressin 1a receptors
Sci. Rep., 9 (1) (2019), p. 15480, 10.1038/s41598-019-52024-9
Google Scholar
Functional New World monkey oxytocin forms elicit an altered signaling profile and promotes parental care in rats
Proc. Natl. Acad. Sci. U. S. A., 114 (34) (2017), pp. 9044-9049, 10.1073/pnas.1711687114
Google Scholar
Oxytocin and vasopressin are dysregulated in Williams Syndrome, a genetic disorder affecting social behavior
PLoS One, 7 (6) (2012), Article e38513, 10.1371/journal.pone.0038513
Google Scholar
Plasma oxytocin explains individual differences in neural substrates of social perception
Front. Hum. Neurosci., 9 (2015), p. 132, 10.3389/fnhum.2015.00132
Google Scholar
Challenges for measuring oxytocin: the blind men and the elephant?
Psychoneuroendocrinology, 107 (2019), pp. 225-231, 10.1016/j.psyneuen.2019.05.018
Google Scholar
Specificity of plasma oxytocin immunoassays: a comparison of commercial assays and sample preparation techniques using oxytocin knockout and wildtype mice
Psychoneuroendocrinology, 132 (2021), p. 105368, 10.1016/j.psyneuen.2021.105368
Google Scholar
An improved sample extraction method reveals that plasma receptor for advanced glycation end-products (RAGE) modulates circulating free oxytocin in mice
Peptides (September 17, 2021), p. 170649, 10.1016/j.peptides.2021.170649
Published online
Google Scholar
Intranasal oxytocin: myths and delusions
Biol. Psychiatr., 79 (3) (2016), pp. 243-250, 10.1016/j.biopsych.2015.05.003
Google Scholar
Oxytocin in the tumor microenvironment is associated with lower inflammation and longer survival in advanced epithelial ovarian cancer patients
Psychoneuroendocrinology, 106 (2019), pp. 244-251, 10.1016/j.psyneuen.2019.04.007
Google Scholar
1465771; on some physiological actions of ergot
J. Physiol., 34 (3) (1906), pp. 163-206
Google Scholar
Brain oxytocin: how puzzle stones from animal studies translate into psychiatry
Mol. Psychiatr., 26 (1) (2021), pp. 265-279, 10.1038/s41380-020-0802-9
Google Scholar
Oxytocin gene expression in rat uterus
Science, 256 (5063) (1992), pp. 1553-1555, 10.1126/science.1598587
Google Scholar
Positive psychosocial factors and oxytocin in the ovarian tumor microenvironment
Psychosom. Med., 83 (5) (2021), pp. 417-422, 10.1097/PSY.0000000000000935
Google Scholar
The oxytocin receptor: from intracellular signaling to behavior
Physiol. Rev., 98 (3) (2018), pp. 1805-1908, 10.1152/physrev.00031.2017
Google Scholar
Anxiolytic and anxiogenic? How the transcription factor MEF2 might explain the manifold behavioral effects of oxytocin
Front. Endocrinol., 11 (2020), p. 186, 10.3389/fendo.2020.00186
Google Scholar
Oxytocin and vasopressin: powerful regulators of social behavior
Neuroscientist, 23 (5) (2017), pp. 517-528, 10.1177/1073858417708284
Google Scholar
Cross-talk among oxytocin and arginine-vasopressin receptors: relevance for basic and clinical studies of the brain and periphery
Front. Neuroendocrinol., 51 (2018), pp. 14-24, 10.1016/j.yfrne.2017.10.004
Google Scholar
Vasopressin stimulates ventromedial hypothalamic neurons via oxytocin receptors in oxytocin gene knockout male and female mice
Neuroendocrinology, 80 (2) (2004), pp. 92-99, 10.1159/000081844
Google Scholar
Astrocytes mediate the effect of oxytocin in the central amygdala on neuronal activity and affective states in rodents
Nat. Neurosci., 24 (4) (2021), pp. 529-541, 10.1038/s41593-021-00800-0
Google Scholar
Crystal structure of the human oxytocin receptor
Sci. Adv., 6 (29) (2020), Article eabb5419, 10.1126/sciadv.abb5419
Google Scholar
Uterine excitability and ion channels and their changes with gestation and hormonal environment
Annu. Rev. Physiol., 83 (2021), pp. 331-357, 10.1146/annurev-physiol-032420-035509
Google Scholar
H2S and oxytocin systems in early life stress and cardiovascular disease
J. Clin. Med., 10 (16) (2021), p. 3484, 10.3390/jcm10163484
Google Scholar
Vasopressin and oxytocin receptor systems in the brain: sex differences and sex-specific regulation of social behavior
Front. Neuroendocrinol., 40 (2016), pp. 1-23, 10.1016/j.yfrne.2015.04.003
Google Scholar
Relation of oxytocin to psychological stress responses and hypothalamic-pituitary-adrenocortical axis activity in older women
Psychosom. Med., 68 (2) (2006), pp. 238-245, 10.1097/01.psy.0000203242.95990.74
Google Scholar
Sex differences in the neural and behavioral response to intranasal oxytocin and vasopressin during human social interaction
Psychoneuroendocrinology, 39 (2014), pp. 237-248, 10.1016/j.psyneuen.2013.09.022
Google Scholar
The monogamy paradox: what do love and sex have to do with it?
Front. Ecol. Evol., 6 (202) (2018), 10.3389/fevo.2018.00202
Google Scholar
Oxytocin and steroid actions
Curr Top Behav Neurosci, 35 (2018), pp. 77-95, pp. 747-755, 10.1007/7854_2017_9
Google Scholar
Beyond infection - maternal immune activation by environmental factors, microglial development, and relevance for autism spectrum disorders
Exp. Neurol., 299 (2018), pp. 241-251, 10.1016/j.expneurol.2017.07.002
Journal Article
Google Scholar
A new view of sexual differentiation of mammalian brain
J. Comp. Physiol. A Neuroethol. Sens. Neural Behav. Physiol., 206 (3) (2020), pp. 369-378, 10.1007/s00359-019-01376-8
Google Scholar
Consequences of early experiences and exposure to oxytocin and vasopressin are sexually dimorphic
Dev. Neurosci., 31 (4) (2009), pp. 332-341
Google Scholar
Regulation of the hypothalamic-pituitary-adrenal axis by neuropeptides
Horm. Mol. Biol. Clin. Invest., 7 (2) (2011), pp. 327-336, 10.1515/HMBCI.2011.123
Google Scholar
Magnocellular vasopressin and the mechanism of “glucocorticoid escape
Front. Endocrinol., 10 (2019), p. 422, 10.3389/fendo.2019.00422
Google Scholar
Sex differences in stress reactivity in arousal and attention systems
Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol., 44 (1) (2019), pp. 129-139, 10.1038/s41386-018-0137-2
Google Scholar
Roles for androgens in mediating the sex differences of neuroendocrine and behavioral stress responses
Biol. Sex Differ., 11 (1) (2020), p. 44, 10.1186/s13293-020-00319-2
Google Scholar
Dissociation of oxytocin, vasopressin and corticotropin secretion during different types of stress
Life Sci., 35 (5) (1984), pp. 487-491
Google Scholar
Oxytocin knockout mice: a model for studying stress-related and ingestive behaviours
Prog. Brain Res., 170 (2008), pp. 53-64, 10.1016/S0079-6123(08)00405-6
Google Scholar
Oxytocin and sexual behavior
Neurosci. Biobehav. Rev., 16 (2) (1992), pp. 131-144, 10.1016/s0149-7634(05)80176-9
Google Scholar
Oxytocin in the male reproductive tract; the therapeutic potential of oxytocin-agonists and-antagonists
Front. Endocrinol., 11 (2020), p. 565731, 10.3389/fendo.2020.565731
Google Scholar
Oxytocin: its mechanism of action and receptor signalling in the myometrium
J. Neuroendocrinol., 26 (6) (2014), pp. 356-369, 10.1111/jne.12154
Google Scholar
The effects of oxytocin and vasopressin on partner preferences in male and female prairie voles (Microtus ochrogaster)
Behav. Neurosci., 113 (5) (1999), pp. 1071-1079
Google Scholar
Salivary oxytocin concentrations in response to running, sexual self-stimulation, breastfeeding and the TSST: the Regensburg Oxytocin Challenge (ROC) study
Psychoneuroendocrinology, 62 (2015), pp. 381-388, 10.1016/j.psyneuen.2015.08.027
Google Scholar
The associations between oxytocin and trauma in humans: a systematic review
Front. Pharmacol., 9 (2018), p. 154, 10.3389/fphar.2018.00154
Google Scholar
The osmoresponsiveness of oxytocin and vasopressin neurones: mechanisms, allostasis and evolution
J. Neuroendocrinol., 31 (3) (2019), Article e12662, 10.1111/jne.12662
Google Scholar
Calming cycle theory and the Co-regulation of oxytocin
Psychodyn. Psychiatry, 45 (4) (2017), pp. 519-540, 10.1521/pdps.2017.45.4.519
Google Scholar
An allostatic theory of oxytocin
Trends Cognit. Sci., 24 (7) (2020), pp. 515-528, 10.1016/j.tics.2020.03.008
Google Scholar
Oxytocin released within the supraoptic nucleus of the rat brain by positive feedback action is involved in parturition-related events
J. Neuroendocrinol., 8 (3) (1996), pp. 227-233
Google Scholar
Peripheral oxytocin administration buffers autonomic but not behavioral responses to environmental stressors in isolated prairie voles
Stress, 15 (2) (2012), pp. 149-161
Google Scholar
Arginine vasopressin in the medial amygdala causes greater post-stress recruitment of hypothalamic vasopressin neurons
Mol. Brain, 14 (1) (2021), p. 141, 10.1186/s13041-021-00850-2
Google Scholar
A novel role of oxytocin: oxytocin-induced well-being in humans
Biophys. Physicobiol., 16 (2019), pp. 132-139, 10.2142/biophysico.16.0_132
Google Scholar
Oxytocin signaling in pain: cellular, circuit, system, and behavioral levels
Curr Top Behav Neurosci, 35 (2018), pp. 193-211, 10.1007/7854_2017_14
Google Scholar
The role of oxytocin, vasopressin, and their receptors at nociceptors in peripheral pain modulation
Front. Neuroendocrinol., 63 (2021), p. 100942, 10.1016/j.yfrne.2021.100942
Google Scholar
Oxytocin-induced analgesia and scratching are mediated by the vasopressin-1A receptor in the mouse
J. Neurosci. Off. J. Soc. Neurosci., 30 (24) (2010), pp. 8274-8284, 10.1523/JNEUROSCI.1594-10.2010
Google Scholar
An epigenetic rheostat of experience: DNA methylation of OXTR as a mechanism of early life allostasis
Compr. Psychoneuroendocrinol., 8 (2021), p. 100098, 10.1016/j.cpnec.2021.100098
Google Scholar
Early nurture epigenetically tunes the oxytocin receptor
Psychoneuroendocrinology, 99 (2019), pp. 128-136, 10.1016/j.psyneuen.2018.08.037
Google Scholar
Behavioral and epigenetic consequences of oxytocin treatment at birth
Sci. Adv., 5 (5) (2019), Article eaav2244, 10.1126/sciadv.aav2244
Google Scholar
Developmental programming of oxytocin through variation in early-life stress: four meta-analyses and a theoretical reinterpretation
Clin. Psychol. Rev., 86 (2021), p. 101985, 10.1016/j.cpr.2021.101985
Google Scholar
Developmental exposure to vasopressin increases aggression in adult prairie voles
Proc. Natl. Acad. Sci. USA, 96 (22) (1999), pp. 12601-12604
Google Scholar
Sex differences in the regulation of social and anxiety-related behaviors: insights from vasopressin and oxytocin brain systems
Curr. Opin. Neurobiol., 49 (2018), pp. 132-140, 10.1016/j.conb.2018.02.011
Google Scholar
Early life adversity and adult social behavior: focus on arginine vasopressin and oxytocin as potential mediators
Front. Behav. Neurosci., 13 (2019), p. 143, 10.3389/fnbeh.2019.00143
Google Scholar
Oxytocin/vasopressin-related peptides have an ancient role in reproductive behavior
Science, 338 (6106) (2012), pp. 540-543, 10.1126/science.1226201
Google Scholar
Assembling the puzzle: pathways of oxytocin signaling in the brain
Biol. Psychiatr., 79 (3) (2016), pp. 155-164, 10.1016/j.biopsych.2015.04.013
Google Scholar
Man and the chimaera. Selective versus neutral oxytocin evolution
Adv. Exp. Med. Biol., 395 (1995), pp. 615-627
Google Scholar
Evolution of oxytocin pathways in the brain of vertebrates
Front. Behav. Neurosci., 8 (2014), p. 31, 10.3389/fnbeh.2014.00031
Google Scholar
Characterization of the neurohypophysial hormone gene loci in elephant shark and the Japanese lamprey: origin of the vertebrate neurohypophysial hormone genes
BMC Evol. Biol., 9 (2009), p. 47, 10.1186/1471-2148-9-47
Google Scholar
Platypus and echidna genomes reveal mammalian biology and evolution
Nature, 592 (7856) (2021), pp. 756-762, 10.1038/s41586-020-03039-0
Google Scholar
Information processing in affective disorders: did an ancient peptide regulating intercellular metabolism become Co-opted for noxious stress sensing?
BioEssays News Rev. Mol. Cell Dev. Biol., 42 (9) (2020), Article e2000039, 10.1002/bies.202000039
Google Scholar
Oxytocinergic feedback circuitries: an anatomical basis for neuromodulation of social behaviors
Front. Neural Circ., 15 (2021), p. 688234, 10.3389/fncir.2021.688234
Google Scholar
A novel role of CD38 and oxytocin as tandem molecular moderators of human social behavior
Neurosci. Biobehav. Rev., 115 (2020), pp. 251-272, 10.1016/j.neubiorev.2020.04.013
Google Scholar
RAGE regulates oxytocin transport into the brain
Commun. Biol., 3 (1) (2020), 10.1038/s42003-020-0799-2
70-70
Google Scholar
Oxytocin promotes accurate fear discrimination and adaptive defensive behaviors
Front. Neurosci., 14 (2020), p. 583878, 10.3389/fnins.2020.583878
Google Scholar
The oxytocin-vasopressin pathway in the context of love and fear
Front. Endocrinol., 8 (2017), p. 356, 10.3389/fendo.2017.00356
Google Scholar
Prosociality as a foundation for intergroup conflict
Curr. Opin. Psychol., 44 (2021), pp. 112-116, 10.1016/j.copsyc.2021.09.002
Google Scholar
Vasopressin but not oxytocin responds to birth stress in infants
Front. Neurosci., 15 (2021), p. 718056, 10.3389/fnins.2021.718056
Google Scholar
Early life abuse moderates the effects of intranasal oxytocin on symptoms of premenstrual dysphoric disorder: preliminary evidence from a placebo-controlled trial
Front. Psychiatr., 9 (2018), p. 547, 10.3389/fpsyt.2018.00547
Google Scholar
Intranasal oxytocin administration improves mood in new mothers with moderate low mood but not in mothers with elevated symptoms of postnatal depression: a randomised controlled trial
J. Affect. Disord. (2022), 10.1016/j.jad.2021.11.062
S0165-0327(21)01292-1
Google Scholar
Oxytocin, neural plasticity, and social behavior
Annu. Rev. Neurosci. (April 6, 2021), 10.1146/annurev-neuro-102320-102847
Published online
Google Scholar
The neuroendocrine basis of social recognition
Front. Neuroendocrinol., 23 (2) (2002), pp. 200-224, 10.1006/frne.2002.0229
Google Scholar
Exogenous effects of oxytocin in five psychiatric disorders: a systematic review, meta-analyses and a personalized approach through the lens of the social salience hypothesis
Neurosci. Biobehav. Rev., 114 (2020), pp. 70-95, 10.1016/j.neubiorev.2020.04.023
Google Scholar
Parenthood, stress, and the brain
Biol. Psychiatr., 70 (9) (2011), pp. 804-805, 10.1016/j.biopsych.2011.09.003
Google Scholar
The role of oxytocin in social bonding, stress regulation and mental health: an update on the moderating effects of context and interindividual differences
Psychoneuroendocrinology, 38 (9) (2013), pp. 1883-1894, 10.1016/j.psyneuen.2013.06.019
Google Scholar
Love and fear in the times of sickness
Compr. Psychoneuroendocrinol., 6 (2021), p. 100032, 10.1016/j.cpnec.2021.100032
Google Scholar
Interplay of oxytocin, vasopressin, and sex hormones in the regulation of social recognition
Behav. Neurosci., 126 (1) (2012), pp. 97-109, 10.1037/a0026464
Google Scholar
Oxytocin neurons enable social transmission of maternal behaviour
Nature, 596 (7873) (2021), pp. 553-557, 10.1038/s41586-021-03814-7
Google Scholar
The magnocellular oxytocin system, the fount of maternity: adaptations in pregnancy
Front. Neuroendocrinol., 24 (1) (2003), pp. 27-61
Google Scholar
Oxytocin and vasopressin, and the GABA developmental shift during labor and birth: friends or foes?
Front. Cell. Neurosci., 12 (2018), p. 254, 10.3389/fncel.2018.00254
Google Scholar
Social reward requires coordinated activity of nucleus accumbens oxytocin and serotonin
Nature, 501 (2013), p. 179, 10.1038/nature12518
Google Scholar
Nucleus accumbens dopamine differentially mediates the formation and maintenance of monogamous pair bonds
Nat. Neurosci., 9 (1) (2005), pp. 133-139, 10.1038/nn1613
Google Scholar
Blocking mu-opioid receptors inhibits social bonding in rituals
Biol. Lett., 16 (10) (2020), p. 20200485, 10.1098/rsbl.2020.0485
Google Scholar
Early parent-child interactions and substance use disorder: an attachment perspective on a biopsychosocial entanglement
Neurosci. Biobehav. Rev., 131 (2021), pp. 560-580, 10.1016/j.neubiorev.2021.09.052
Google Scholar
The science of love: state of the art
Adv. Exp. Med. Biol., 1331 (2021), pp. 249-254, 10.1007/978-3-030-74046-7_16
Google Scholar
Neuroendocrine perspectives on social attachment and love
Psychoneuroendocrinology, 23 (8) (1998), pp. 779-818
Google Scholar
Beyond “oxytocin = good”: neural complexities and the flipside of social bonds
Arch. Sex. Behav., 42 (7) (2013), pp. 1115-1118, 10.1007/s10508-013-0134-9
Google Scholar
Mother love: what turns it on?
Am. Sci., 59 (4) (1971), pp. 404-407
Google Scholar
Sex, parturition and motherhood without oxytocin?
J. Endocrinol., 157 (3) (1998), pp. 343-359, 10.1677/joe.0.1570343
Google Scholar
Oxytocin is required for nursing but is not essential for parturition or reproductive behavior
Proc. Natl. Acad. Sci. Unit. States Am., 93 (21) (1996), pp. 11699-11704
Google Scholar
Deficiency in mouse oxytocin prevents milk ejection, but not fertility or parturition
J. Neuroendocrinol., 8 (11) (1996), pp. 847-853, 10.1046/j.1365-2826.1996.05266.x
Google Scholar
Olfactory regulation of maternal behavior in rats. II. Effects of peripherally induced anosmia and lesions of the lateral olfactory tract in pup-induced virgins
J. Comp. Physiol. Psychol., 86 (2) (1974), pp. 233-246, 10.1037/h0035936
Google Scholar
Integrative functions of lactational hormones in social behavior and stress management
Ann. N. Y. Acad. Sci., 807 (1997), pp. 164-174
Google Scholar
Elevation of oxytocin levels early post partum in women
Acta Obstet. Gynecol. Scand., 74 (7) (1995), pp. 530-533, 10.3109/00016349509024384
Google Scholar
Sensory stimulation of oxytocin release is associated with stress management and maternal care
Front. Psychol., 11 (2020), p. 588068, 10.3389/fpsyg.2020.588068
Google Scholar
Oxytocin increases the social salience of the outgroup in potential threat contexts
Horm. Behav., 122 (2020), p. 104733, 10.1016/j.yhbeh.2020.104733
Google Scholar
Passionate love/sexual desire: can the same paradigm explain both?
Arch. Sex. Behav., 16 (3) (1987), pp. 259-278, 10.1007/BF01541613
Google Scholar
Intense, passionate, romantic love: a natural addiction? How the fields that investigate romance and substance abuse can inform each other
Front. Psychol., 7 (2016), p. 687, 10.3389/fpsyg.2016.00687
Google Scholar
Metaphor: bridging embodiment to abstraction
Psychon. Bull. Rev., 23 (4) (2016), pp. 1080-1089, 10.3758/s13423-015-0861-0
Google Scholar
Dynamic grounding of emotion concepts
Philos. Trans. R. Soc. Lond. B Biol. Sci., 373 (1752) (2018), p. 20170127, 10.1098/rstb.2017.0127
Google Scholar
Protective environments and health status: cross-talk between human and animal studies
Neurobiol. Aging, 26 (Suppl 1) (2005), pp. 113-118, 10.1016/j.neurobiolaging.2005.08.020
Google Scholar
Polyvagal Theory: a biobehavioral journey to sociality
Compr. Psychoneuroendocrinol., 7 (2021), p. 100069, 10.1016/j.cpnec.2021.100069
Google Scholar
IInterplay between oxytocin and sensory systems in the orchestration of socio-emotional behaviors
Neuron, 99 (5) (2018), pp. 887-904, 10.1016/j.neuron.2018.07.016
Google Scholar
The polyvagal theory: phylogenetic substrates of a social nervous system
Int. J. Psychophysiol., 42 (2) (2001), pp. 123-146, 10.1016/S0167-8760(01)00162-3
Google Scholar
Attachment theory and its therapeutic implications
Adolesc. Psychiatr., 6 (1978), pp. 5-33
Google Scholar
Mother brain is wired for social moments
Elife, 10 (2021), Article e59436, 10.7554/eLife.59436
Google Scholar
Responses to laboratory psychosocial stress in postpartum women
Psychosom. Med., 63 (5) (2001), pp. 814-821, 10.1097/00006842-200109000-00015
Google Scholar
Birth signalling hormones and the developmental consequences of caesarean delivery
J. Neuroendocrinol. (2020), Article e12912, 10.1111/jne.12912
n/a(n/a)
Google Scholar
Early-life oxytocin attenuates the social deficits induced by caesarean-section delivery in the mouse
Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol. (2021), 10.1038/s41386-021-01040-3
Published online May 26
Google Scholar
Developmental perspectives on oxytocin and vasopressin
Neuropsychopharmacology, 40 (1) (2015), pp. 24-42, 10.1038/npp.2014.120
Google Scholar
A learning theory of attachment: unraveling the black box of attachment development
Neurosci. Biobehav. Rev., 113 (2020), pp. 287-298, 10.1016/j.neubiorev.2020.03.014
Google Scholar
A learning theory of attachment: unraveling the black box of attachment development
Neurosci. Biobehav. Rev., 113 (2020), pp. 287-298, 10.1016/j.neubiorev.2020.03.014
Google Scholar
Oxytocin facilitates adaptive fear and attenuates anxiety responses in animal models and human studies-potential interaction with the corticotropin-releasing factor (CRF) system in the bed nucleus of the stria terminalis (BNST)
Cell Tissue Res., 375 (1) (2019), pp. 143-172, 10.1007/s00441-018-2889-8
Google Scholar
Social behavior as a transdiagnostic marker of resilience
Annu. Rev. Clin. Psychol., 17 (2021), pp. 153-180, 10.1146/annurev-clinpsy-081219-102046
Google Scholar
Predictive regulation and human design
Elife, 7 (2018), 10.7554/eLife.36133
Google Scholar
Love, fear, and the human-animal bond: on adversity and multispecies relationships
Compr. Psychoneuroendocrinol., 7 (2021), p. 100071, 10.1016/j.cpnec.2021.100071
Published online
Google Scholar
Fear, love, and the origins of canid domestication: an oxytocin hypothesis
Compr. Psychoneuroendocrinol., 9 (2022), p. 100100, 10.1016/j.cpnec.2021.100100
Google Scholar
Lost connections: oxytocin and the neural, physiological, and behavioral consequences of disrupted relationships
Int. J. Psychophysiol. (January 9, 2018), 10.1016/j.ijpsycho.2017.12.011
Published online
Google Scholar
Loneliness across phylogeny and a call for comparative studies and animal models
Perspect. Psychol. Sci. J. Assoc. Psychol. Sci., 10 (2) (2015), pp. 202-212, 10.1177/1745691614564876
Google Scholar
The role of the cholinergic anti-inflammatory pathway in autoimmune rheumatic diseases
Scand. J. Immunol., 94 (4) (2021), Article e13092, 10.1111/sji.13092
Google Scholar
Oxytocin promotes functional coupling between paraventricular nucleus and both sympathetic and parasympathetic cardioregulatory nuclei
Horm. Behav., 80 (2016), pp. 82-91, 10.1016/j.yhbeh.2016.01.010
Google Scholar
Chronic inflammation in the etiology of disease across the life span
Nat. Med., 25 (12) (2019), pp. 1822-1832, 10.1038/s41591-019-0675-0
Google Scholar
Oxytocin may have a therapeutical potential against cardiovascular disease. Possible pharmaceutical and behavioral approaches
Med. Hypotheses, 138 (2020), p. 109597, 10.1016/j.mehy.2020.109597
Google Scholar
Oxytocin attenuates atherosclerosis and adipose tissue inflammation in socially isolated ApoE-/- mice
Psychosom. Med., 72 (4) (2010), pp. 376-382
Google Scholar
Perspective: cell danger response Biology-The new science that connects environmental health with mitochondria and the rising tide of chronic illness
Mitochondrion, 51 (2020), pp. 40-45, 10.1016/j.mito.2019.12.005
Google Scholar
Oxytocin's anti-inflammatory and proimmune functions in COVID-19: a transcriptomic signature-based approach
Physiol. Genom., 52 (9) (2020), pp. 401-407, 10.1152/physiolgenomics.00095.2020
Google Scholar
Love and peace across generations: biobehavioral systems and global partnerships
Compr. Psychoneuroendocrinol., 8 (2021), p. 100092, 10.1016/j.cpnec.2021.100092
Google Scholar
Oxytocin administration prevents cellular aging caused by social isolation
Psychoneuroendocrinology, 103 (2019), pp. 52-60, 10.1016/j.psyneuen.2019.01.006
Google Scholar
Love and longevity: a social dependency hypothesis
Compr. Psychoneuroendocrinol., 8 (2021), p. 100088, 10.1016/j.cpnec.2021.100088
Google Scholar